The evolution of reproductive modes and life cycles in amphibians - Nature.com
Abstract
Amphibians have undergone important evolutionary transitions in reproductive modes and life-cycles. We compare large-scale macroevolutionary patterns in these transitions across the three major amphibian clades: frogs, salamanders, and caecilians. We analyse matching reproductive and phylogenetic data for 4025 species. We find that having aquatic larvae is ancestral for all three groups and is retained by many extant species (33–44%). The most frequent transitions in each group are to relatively uncommon states: live-bearing in caecilians, paedomorphosis in salamanders, and semi-terrestriality in frogs. All three groups show transitions to more terrestrial reproductive modes, but only in caecilians have these evolved sequentially from most-to-least aquatic. Diversification rates are largely independent of reproductive modes. However, in salamanders direct development accelerates diversification whereas paedomorphosis decreases it. Overall, we find a widespread retention of ancestral modes, decoupling of trait transition rates from patterns of species richness, and the general independence of reproductive modes and diversification.
Introduction
The majority of animals have complex life cycles1, consisting of one or more larval phases and an adult phase (typically with distinct morphologies and ecologies), separated by extensive remodeling of body plans (metamorphosis). However, major clades (e.g., amniotes), have lost the larval stage and exhibit a uniphasic life cycle2. How this loss comes about is an important evolutionary question. This loss has occurred frequently in non-amniote vertebrates, especially Amphibia. Since their origin ~300 million years ago3, amphibians have evolved many alternatives to their ancestral biphasic life cycle4,5, including loss of larval stages (through direct development or viviparity) and loss of the adult stage (paedomorphosis). Here we use a phylogenetic approach to test hypotheses about the evolution of reproductive modes and life cycles among the three major amphibian clades (frogs [Anura], salamanders [Caudata], and caecilians [Gymnophiona]) and their possible consequences for lineage diversification rates. More broadly, we evaluate whether large-scale patterns of reproductive-mode and life-cycle evolution are similar or different among major clades (and how).
Amphibians are an excellent system for comparing life-cycle evolution across major groups. For all three groups, the ancestral reproductive mode is thought to include aquatic larvae5,6, which is retained by numerous species in all groups6 (Fig. 1). All three groups also have many species lacking the larval stage (direct development) and some species in which females give birth to live young7 (viviparity; Fig. 1). However, these three clades also differ in many respects, including body form (tailless frogs, limbless caecilians, and tailed and limbed salamanders), species richness (~7100 frogs, ~700 salamanders, ~200 caecilians), and overall geographic distributions (caecilians are mostly tropical, most salamander families are temperate, and frogs are distributed globally)8,9. These differences extend to reproductive modes as well. Bearing live young is rare in frogs and salamanders but more common in caecilians7. Paedomorphism, defined very narrowly here as the absence of a distinct postmetamorphic adult stage, is absent in frogs and caecilians but occurs in most salamander families10. Life cycles with aquatic eggs have been completely lost in caecilians4. Frogs have greater diversity of reproductive modes than the other groups4.
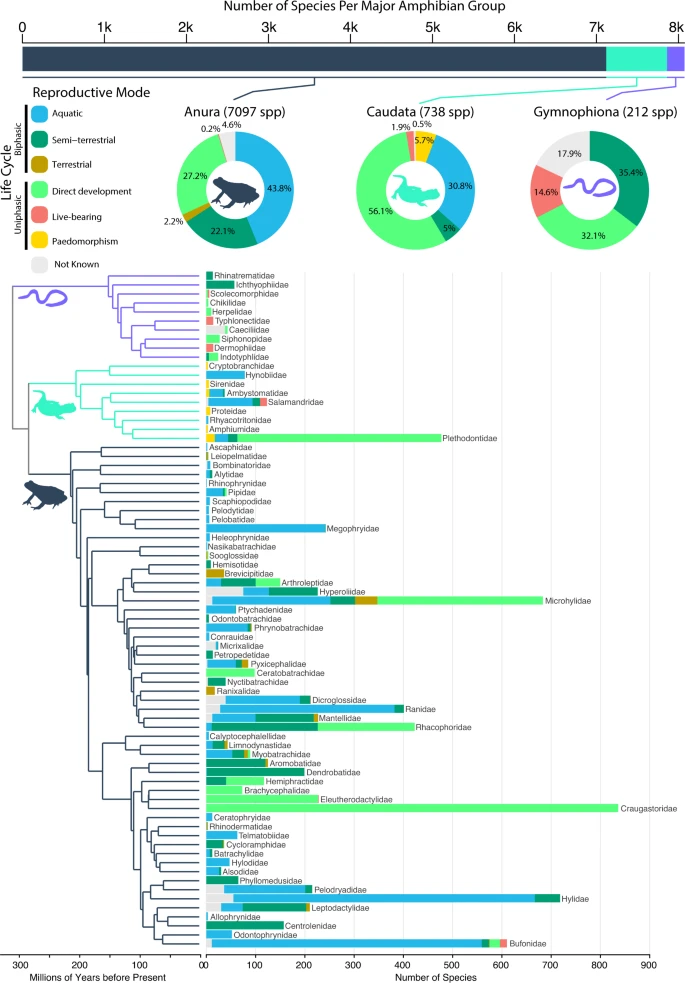
Species richness per amphibian group (top bar chart where Anura: dark blue gray, Caudata: mint green, Gymnophiona: lilac, and 1k = 1000 species) and frequencies of reproductive modes per group (percentages as pie charts), and per family (numbers of species as stacked bars on phylogeny). Reproductive modes are represented as blue = aquatic; dark green = semi-terrestrial; brown = terrestrial; light green = direct development; red = live-bearing; yellow = paedomorphism. Frequencies and percentages of unknown reproductive modes are depicted in light gray. The phylogeny shows the relationships of all 75 amphibian families, with icons and branch colors highlighting the three different amphibian clades. Source data are provided as a Source Data file.
Much diversity in amphibian reproductive modes involves modifications in their life cycles, such as the presence or absence of whole life stages, and whether embryonic and larval development occur in water or on land11,12,13. These two aspects are often tightly linked. For example, most oviparous species that have lost the larval stage (direct development) lay terrestrial eggs4. Amphibian reproductive modes are traditionally thought to have evolved sequentially with only single ontogenetic modules (life stages) changing at each time4,14,15. Following this view (Fig. 2), the ancestral mode is biphasic, fully aquatic development16. The first step is the evolution of a terrestrial egg, but maintaining an aquatic larval stage, followed by a terrestrial larval stage, before losing the larval stage to become direct developing (i.e., post-metamorphic juveniles hatching from eggs). Live-bearing is the most derived mode, in which embryos are retained (with or without the egg) in the oviduct of the mother until metamorphosis or late stages of larval development.

Null: transitions between all states (reproductive modes) can occur. Non-sequential: derived (more terrestrial) states can only transition to and from the ancestral aquatic mode. Sequential: transitions can only occur between the ecologically most similar states. Semi-sequential: derived, fully terrestrial modes (direct development, live bearing and fully terrestrial biphasic) can only transition to and from the semi-terrestrial mode. Up to three variations of this final scenario were tested (dashed lines), allowing live-bearing to transition either to and from the aquatic mode, the semi-terrestrial mode, or direct development. Node colors and annotations reflect reproductive modes, where A, blue = aquatic; S, dark green = semi-terrestrial; T, brown = terrestrial; D, light green = direct development; L red = live-bearing; P, yellow = paedomorphism. Network edge colors and icons distinguish the three amphibian groups, where Anura: dark blue-gray, Caudata: mint green, Gymnophiona: lilac. Because not all six states are present in all three major clades of amphibians (nodes in gray), the scenarios vary between them.
The sequential hypothesis has been partially tested for several amphibian clades. Transitions to more terrestrial modes appear to have occurred sequentially in some clades (e.g., rhacophorid17 and afrobatrachian18 frogs, caecilians19), but not others (e.g., leptodactylid frogs20). Most importantly, a study of 720 frog species (~10% of anurans) suggested that direct development may have evolved directly from fully aquatic ancestors as frequently as from intermediate semi-terrestrial modes21. However, this hypothesis has not been tested across all major groups of amphibians. Moreover, few alternatives to strictly sequential versus strictly non-sequential scenarios (where derived traits all evolve from the ancestral state directly) have been explored. For example, evolving a terrestrially adapted egg may be an evolutionary precursor for completely terrestrial life cycles with terrestrial eggs (e.g., direct development), but not for life cycles without eggs (e.g., live-bearing). Scenarios where only modes that have shared features (e.g., terrestrial eggs) have evolved sequentially therefore need to be considered. There may also be reversals from more terrestrial modes to more aquatic modes21, which would be inconsistent with the traditional sequential scenario of increasing terrestriality.
We also need to understand how transitions among reproductive modes and life cycles may have impacted diversification. The observation that some of the most species-rich amphibian clades have direct development has led to speculation that this mode may drive increased diversification rates22,23,24,25. Diversification rate is the rate of species accumulation over time, or the speciation rate minus the extinction rate. The independence of direct-developing lineages from aquatic habitats may have facilitated their expansion into new niche space (e.g., arboreal microhabitats in frogs and salamanders). Occupation of new niche space can lead to ecological opportunity26 and thus to increased diversification rates through reduced extinction and/or increased speciation. Furthermore, increased terrestriality is generally accompanied by increased egg size, decreased clutch size, reduced body size21, and increased parental care21,27,28,29. These changes, especially reduced body size, may reduce dispersal ability and thereby increase diversification, by increasing allopatric speciation30,31. However, reduced dispersal ability might decrease long-term diversification instead32, since smaller clade-level range sizes might decrease opportunities for allopatric speciation and make species more susceptible to extinction. The idea that reproductive mode influences diversification has seldom been tested in amphibians at a broad scale. Relevant studies have provided evidence for this hypothesis (e.g., Rhacophoridae33) and against it (e.g., Phrynobatrachus34 and phytotelma-breeding frogs35). A study across frogs found that reproductive modes did not significantly impact diversification, but had limited species-level sampling21. Two studies in salamanders found that paedomorphosis significantly decreases diversification rates36,37, with one study on plethodontids37 linking this pattern to reduced geographic range sizes in paedomorphic species (which have limited ability to disperse overland). In summary, there is some equivocal evidence that terrestrial reproduction might increase diversification in frogs and salamanders, and some evidence that changes in life cycle through paedomorphosis decreases diversification in salamanders, but large-scale tests have been limited overall.
There is extensive literature on amphibian reproductive-mode evolution, but core aspects remain untested across the major amphibian groups. Here, we address three open, long-standing questions by taking advantage of a relatively complete, amphibian-wide phylogeny, advanced trait-evolution models, and a growing knowledgebase on amphibian reproductive modes. First, how frequent are different reproductive modes among species within and among major amphibian clades (frogs, salamanders, and caecilians)? Second, do reproductive modes evolve sequentially, from more aquatic to more terrestrial, or in a more complex way? Third, do different reproductive modes or life cycles differentially impact rates of diversification? More broadly, we ask whether these patterns of reproductive-mode evolution are shared or differ among these groups.
Results
Distribution of reproductive modes across amphibians
We performed a literature survey and coded reproductive modes for 95% of described, extant amphibian species (7681 of 8047; Supplementary Data 1). These included 95.4% of anuran species, 99.5% of caudates, and 82.1% of gymnophionans. We categorized each species into one of six modes (Fig. 1): aquatic (aquatic eggs and larva), semi-terrestrial (terrestrial eggs, aquatic larva), terrestrial (terrestrial eggs and larva), direct developing (terrestrial eggs, no larva), live-bearing (no eggs, no larva), or paedomorphic (no adult stage).
Surprisingly, the majority of sampled amphibians lay eggs on land, not water (Anura = 51.5% of n = 7097 sampled species in total; Caudata = 61.1% of n = 738 species; Gymnophiona = 67.5% of n = 212 species; Fig. 1). In anurans, biphasic fully aquatic reproduction (eggs and larvae develop in water) was the most common mode, present in 43.8% of sampled species. In contrast, direct development was the most common mode in Caudata (56.1% of species), but was present in only 27.2% of Anura. In Gymnophiona, the most common states were semi-terrestrial biphasic (35.4%; eggs on land, but larvae develop in water) and direct development (32.1%). Semi-terrestrial, biphasic reproduction was also widespread in Anura (20.9%), but not Caudata (5%). Obligate paedomorphism was present only in Caudata, and was relatively rare (5.7%). Live-bearing was rare in Anura (0.2%) and Caudata (1.9%), and more common in Gymnophiona (14.6%).
Reproductive-mode evolution
We confirmed that the most recent common ancestor of amphibians most likely had aquatic larvae that hatched from aquatic eggs (Supplementary Figs. 7 and 9). We then tested four scenarios for how more terrestrial modes evolved within each group, by constraining transition rates (Fig. 2). These models were: (1) transitions between all modes are possible (null scenario); (2) all derived modes evolve directly and independently from a fully aquatic ancestral state (non-sequential scenario); (3) modes evolve sequentially from most aquatic to least aquatic (sequential scenario); and (4) semi-terrestriality (terrestrial eggs with aquatic larvae) precedes other modes with terrestrial eggs (fully terrestrial and direct development), but transitions are otherwise unordered (semi-sequential scenario). For all models, transitions were allowed to be bidirectional. We compared the fit (using Akaike Information Criterion [AIC]38 and Akaike weights [AICw]39) among scenarios by fitting hidden Markov models of discrete character evolution with the R package corHMM40. We tested different rate restrictions (equal, symmetric or all-rates-different; see Methods) and the inclusion of hidden states to allow for rate variation within observed states. This resulted in up to 36 models per group (full model-fitting results in Supplementary Tables 1–4).
The best-fitting scenario for Anura was semi-sequential (AICw = 0.732; Supplementary Table 1 and Fig. 3). Here, the semi-terrestrial mode evolved from the ancestral aquatic mode, and gave rise to the other two modes with terrestrial eggs: fully terrestrial and direct development (semi-sequential scenario 1 in Fig. 2). Live-bearing evolved directly from aquatic ancestry. This model also allowed for different rates across transitions (all-rates-different: ARD) and within-state transition-rate variation, modeled as hidden states. One other model had AICw>0.01 (AICw = 0.267, ΔAIC = 2.019; Supplementary Table 1). This model differed in that live-bearing evolved from the semi-terrestrial mode, not the aquatic mode (semi-sequential scenario 2 in Fig. 2).

a Estimated transition rates (×10−2) from best-performing corHMM models. Gray arrows and annotations represent hidden state rates. Rates lower than 0.01 are not shown. The best models for Anura and Caudata allowed for all rates to be different (except for pairs of rates annotated with an asterisk that have been forced to be symmetrical; see Methods), and the model for Gymnophiona had all rates set to be equal. b Numbers of transitions when estimating ancestral states at nodes only using joint estimations [Node-to-Node] and the mean numbers of transitions along branches [Anagenetic], estimated from 1000 stochastic character maps. Dashed lines indicate transitions only recovered using one of the two methods. The number of species per reproductive mode represented in the phylogeny are indicated in the nodes of the networks. c Proportions of models in the posterior where a given transition was estimated (i.e., non-zero transition rate) by the reverse jump MCMC of the MultiState Covarion algorithm implemented in BayesTraits. Proportions of 1 indicate transitions that were present in all models sampled by the MCMC. Transitions with proportions <0.5 are not shown. In all cases, color, position and letters of nodes are consistent and represent A, blue = aquatic; S, dark green = semi-terrestrial; T, brown = terrestrial; D, light green = direct development; L red = live-bearing; P, yellow = paedomorphism. Arrow weight is roughly proportional to transition rates (not to scale). Icons for each panel row distinguish the three amphibian groups. Source data are provided as a Source Data file.
The same scenario (semi-sequential scenario; Fig. 2) also had the best fit in Caudata (AICw = 0.454; Fig. 3). For Caudata, nine models fell within AICw > 0.01 and two with ΔAIC < 2 relative to the best-fit model (Supplementary Table 2). Of these nine models, seven were variants of the same scenario but with different rate restrictions and with live-bearing evolving directly from the aquatic or semi-terrestrial states (semi-sequential scenarios 1 or 2 in Fig. 2).
For Gymnophiona, a purely sequential scenario (sequential scenario in Fig. 2) with no hidden states and equal transition rates was the top-ranking model (AICw = 0.421; Fig. 3). A variant of this scenario with symmetrical rates fell within ΔAIC < 2 (ΔAIC = 1.88, AICw = 0.164; Supplementary Table 3) and 11 models had AICw > 0.01.
To test if we overlooked important alternative models, we also conducted a hypothesis-free, model-reduction analysis using the MultiState Reversible-Jump Markov chain Monte Carlo (rjMCMC) implementation of BayesTraits V441. This method reduces the model complexity of the full transition-rate matrix ("null scenario" in Fig. 2), by restricting some transition rates to be zero or equal to each other. These simpler transition models are then sampled in direct proportion to their fit in the MCMC42. Transitions that are not well supported by the data are therefore more frequently set to zero in the posterior. The proportions of times the different transitions were sampled in the posterior (i.e., non-zero) resulted in transition models that was largely congruent with the best-performing corHMM models for each group (Fig. 3c). However, for Anura transitions from fully aquatic to both the fully terrestrial mode and direct development were also supported (present in 100% of the posterior), as were transitions from direct development to the semi-terrestrial mode and from the terrestrial mode to direct development. This last transition was only present in 54% of the posterior, however. For Caudata, the BayesTraits analysis supported transitions from direct development to the aquatic mode and transitions from semi-terrestrial to live-bearing. The latter was represented in 65% of the posterior and therefore arguably ambiguous. Nonetheless, no transition from aquatic to live-bearing was supported. Thus, the transition scenario from BayesTraits was more similar to the second-best corHMM model where this transition was also recovered (Supplementary Table 2; semi-sequential scenario 2 in Fig. 2).
Rates and frequencies of reproductive-mode transitions
The best-performing model for Gymnophiona had identical transition rates fixed between states. For Anura and Caudata, the best-performing models had variable rates, with the highest transition rates away from the semi-terrestrial mode (Fig. 3a). For Anura and Caudata, rates to and from live-bearing (fixed to be symmetrical due to methodological limitations; see Methods) were very low. The lowest rate in Caudata was from paedomorphism to aquatic and in Anura from direct development to semi-terrestrial (all rates <0.001 events/million years [Myr]).
We used these best-fit models (Supplementary Tables 1–3) to estimate frequencies of reproductive-mode transitions across trees (Fig. 3b). We applied two approaches. We used joint ancestral-state estimates to count only transitions between nodes, and stochastic-character mapping (SCM) to count mean anagenetic transitions. The latter can capture multiple transitions along a single branch (see phylogenies with reconstructed states in Supplementary Figs. 1 and 2). In both cases, frequencies are contingent on the model used to estimate them. For Caudata and Gymnophiona, the two methods result in largely congruent estimates of transition frequencies and patterns, but for Anura important discordances exist (Fig. 3b).
For Anura (Fig. 3b), regardless of the method used, semi-terrestriality evolved most frequently (114 times from joint estimates; 144.6 from SCM), followed by direct development (32 versus 37.7). Live-bearing evolved least frequently (2 times). The joint estimates suggested that the complete terrestriality and direct development sometimes evolved directly from the ancestral, aquatic biphasic mode (8 and 3 times, respectively), but these transitions were not found using SCM.
In Caudata (Fig. 3b), paedomorphism evolved most frequently (10 times, both methods), and always from the aquatic mode. Semi-terrestriality evolved many times (joint estimation = 6; SCM = 7.7), mostly from an aquatic ancestor, but with one reversal from direct development. The terrestrial egg was lost frequently through reversals to the fully aquatic mode (joint estimation = 6; SCM = 8.2). Direct development evolved three times, always from a semi-terrestrial ancestor, with one estimated reversal (loss). Live-bearing evolved only once, from an aquatic ancestor.
In Gymnophiona (Fig. 3b), the most frequent transitions were from direct development to live-bearing (joint estimation = 3; SCM = 3.64). Reversals from direct development to semi-terrestrial were twice as common as origins of direct development. There were no reversals from live-bearing.
State-dependent diversification
We used extensions of the State-dependent Speciation and Extinction (SSE) framework43,44 to test whether different reproductive modes or life cycles were associated with different diversification rates (speciation minus extinction). We used the hisse45 and secsse46 R packages, which both allow for multi-state and hidden-state modeling (but each with limitations, see Methods). We used hisse to test whether rate shifts occurred in association with shifts in life cycle, specifically the loss of the larval phase (direct development and live-bearing) or adult phase (paedomorphism). We used secsse to test rate shifts associated with reproductive modes (five states for Anura and Caudata, three for Gymnophiona). For both analyses we tested the fit (based on AIC and AICw) of three scenarios: (1) diversification rates are constant across the phylogeny; (2) diversification rates vary with reproductive mode (with or without rate variation across hidden states); and (3) diversification rates vary, but only across unmeasured (hidden) trait states, not across the observed reproductive modes. The parameters for the best-fit models for all three groups are provided as Supplementary Data 2.
Diversification rates varied across the phylogeny in Anura and Caudata, but not Gymnophiona. This rate variation was attributable to shifts in reproductive modes in Caudata, but not Anura. In anurans, variable rates were due to other, unmeasured (i.e., hidden) traits (Tables 1 and 2). This was the case regardless of the method and trait classification used (hisse with up to three life cycles, and secsse with up to five reproductive modes).
For anurans, the state-independent (variable) models outperformed all others (hisse: AICw = 1.000; secsse: AICw = 0.852; Tables 1 and 2). The secsse analysis estimated a diversification rate of 0.080 species/Myr across all reproductive modes when averaging across hidden and observed state rates (Fig. 4b and Supporting Data 2). Estimating tip (i.e., extant species) net diversification rates using the hisse model yielded similar rate distributions for biphasic and direct-developing species, centered on mean rates of 0.058 species/Myrs for both (sd = 0.016 and sd = 0.013 respectively; Fig. 4b).

a Distributions (with standard deviation error bars around the mean) of rates estimated per species in the phylogeny for each life-cycle category (i.e., tip rates). Species are categorized into biphasic life cycles (dark magenta), direct development (dark blue) or paedomorphism (yellow). Sample sizes (where n = number of species) for each are Anura: biphasic = 2608, direct development = 755; Caudata: biphasic = 192, direct development = 320, paedomorphism = 34; Gymnophiona: biphasic = 19, direct development = 43. Estimates were calculated using the best-fitting hisse models, which for Anura was a Reproductive mode Independent Diversification rate model (RmID) with hidden states, for Caudata was a Reproductive mode dependent Diversification rate model (RmDD) with hidden states, and for Gymnophiona was a Constant Diversification rate model (CD). In cases where models contained hidden states, rates are weighted averages per species. b Diversification rate estimates for each of the six reproductive modes, where blue = aquatic; dark green = semi-terrestrial; brown = terrestrial; light green = direct development; red = live-bearing; yellow = paedomorphism. Estimates are calculated using the best-fitting secsse models, which for Anura was a Reproductive mode Independent Diversification rate model (RmID) with hidden states, for Caudata was a Reproductive mode dependent Diversification model (RmDD) with hidden states, and for Gymnophiona was a Constant Diversification rate model (CD). In cases where models contained hidden states, rates shown are averaged across hidden and observed states. Icons in both panels distinguish the three amphibian groups. Source data are provided as a Source Data file.
In Caudata, the best-performing models were reproductive-mode dependent models with hidden states, for both hisse and secsse (AICw = 0.990 and AICw = 1.000 respectively; Tables 1 and 2). When estimating tip net-diversification rates with this best-performing hisse model, direct-developing caudates had the highest mean net-diversification rates (mean = 0.113 species/Myr, sd = 0.023; Fig. 4a). Biphasic species had intermediate rates (mean = 0.065, sd = 0.034), and paedomorphic species had the lowest rates (mean = −0.022, sd = 0.010). Rate distributions were generally multi-modal or skewed (due to variation in rates across hidden-trait states), and therefore mean rates should be interpreted with caution. The secsse model estimated the highest rates (averaged across observed and hidden states) for direct development (0.094 lineages/Myr) followed by aquatic (0.044), semi-terrestrial (0.043), live-bearing (0.019), and paedomorphism (−0.010; Fig. 4b and Supporting Data 2).
The best-performing models for Gymnophiona were constant-rate models for both hisse and secsse (hisse: AICw = 0.809; AICw = secsse: 0.900; Tables 1 and 2). All species were therefore estimated to diversify at 0.031 species/Myr with hisse, and 0.029 with secsse.
Discussion
Here we provide a large-scale phylogenetic comparison of macroevolutionary patterns in life-cycle and reproductive-mode evolution among the three major groups of amphibians. We specifically focused on comparing species richness of different reproductive modes, testing alternative evolutionary scenarios of transitions among modes, and variation in diversification rates associated with each mode.
Frequencies and transitions of reproductive modes
Our compilation of reproductive modes, the largest for amphibians to date, shows that the three groups have different patterns of species richness among reproductive modes and life cycles (Fig. 1). Frogs and salamanders have many species with fully aquatic reproductive modes, whereas caecilians have none. Salamanders have paedomorphic species, whereas the other two groups do not. Salamanders are dominated by species with direct development and have very few semi-terrestrial species: both frogs and caecilians have similar proportions of species with direct development and with semi-terrestrial reproduction. Caecilians have many live-bearing species, whereas salamanders and frogs have very few.
At the same time, there are important similarities among groups in these richness patterns. For example, all three groups have many species with direct development. All three groups have many species retaining the most ancestral mode. No groups are dominated by live-bearing (Anura: 0.2%, Caudata: 1.9%, Gymnophiona: 14.6%) or paedomorphic species (5.7% in Caudata, 0% in all others). This is particularly interesting given that in caudates the most frequent transitions are gains of paedomorphism, and in gymnophionans gains of live-bearing.
Our ancestral-state reconstructions found that the most recent common ancestors for all three amphibian groups had biphasic life cycles that were subsequently lost multiple times. Alternative larval stages such as non-feeding lecithotrophy, have evolved multiple times across metazoans47,48, and evolutionary losses of larval stages have occurred frequently49,50. The re-evolution of lost, complex traits (such as entire life-stages) was long thought to be impossible. However, a growing number of studies suggest otherwise51,52. The re-appearance of the larval stage after its loss may have occurred in frogs53, salamanders54 and caecilians19. In line with these studies, we found support for reversals from direct development in salamanders and caecilians, with some support in frogs (in the BayesTraits analysis). The detection of reversals can be influenced by the models and methods used for ancestral-state estimations and by the taxon sampling and phylogenetic trees used53,55. For example, previous studies found re-appearance of the larval stage in hemiphractid frogs53,55, which we did not recover with our ancestral-state reconstructions based on corHMM models. This is possibly because rate heterogeneity within hemiphractids may make this pattern difficult to detect53, a problem that would be exacerbated when analyzing all frogs simultaneously. The re-appearance of the larval stage in Anura, Caudata, and Gymnophiona is therefore possible, but rare. In all three groups, transition frequencies are exceedingly asymmetrical, favouring loss of the larval stage. The ontogeny of different direct-developing lineages56,57 may explain why so few direct-developing clades have had reversals. For example, direct-developing hemiphractid frogs have retained some biphasic embryonic features (e.g., oral structures) inside the egg53,58. The same is true for the reversal in Desmognathus salamanders59 and Seychellean caecilians60. Reversals may therefore be viable only if key larval features were retained. Moreover, there is discussion over whether these apparent reversals represent true "re-evolution" of larval stages or further elaborations of existing developmental sequences59. Studies on the regulatory genomic changes required to evolve direct development or live-bearing may therefore be crucial for understanding how these transitions occur and why they are seldom reversed61,62.
Reproductive-mode evolution
We used both hypothesis-driven (corHMM) and hypothesis-free (BayesTraits) approaches to investigate reproductive-mode evolution in amphibians. Caecilians were the only group in which evolution proceeded sequentially from most aquatic to least aquatic (semi-terrestrial to direct developing to live-bearing), as predicted4. In Anura and Caudata, the sequential models were not among the best-fitting scenarios (AICw ≤ 0.01; Supplementary Tables 1 and 2). Lineages with terrestrial larvae did not give rise to direct-developing lineages (transition absent in the best-fit corHMM model and only weakly supported by BayesTraits) and direct developers did not evolve live-bearing. Instead, these three modes represent end-points of different trajectories for evolving terrestrial life cycles. Congruently, although all three modes are ecologically similar (minimal dependency on aquatic habitats), they have diverged extensively in their developmental biology15,63.
A crucial sequential element is maintained in all three groups: the semi-terrestrial mode appears to have been a frequent precursor for direct-development. Our best-fit, semi-sequential hypothesis is therefore congruent with previous reports of sequential evolution in the frog clades Rhacophoridae17,33 and Afrobatrachia18. Neither of these clades have live-bearing species and they show only that terrestrial eggs evolved prior to terrestrial (or late hatching) larvae. Our hypothesis is also congruent with the reported non-sequential evolution in Leptodactylidae20. This study used a more fine-grained categorization of reproductive modes (e.g., distinguishing between foam and non-foam terrestrial nests), in species that fall within only two of our reproductive categories: semi-terrestrial and terrestrial. As such, our more generalized, semi-sequential model also adequately describes reproductive-mode transitions in this group.
Some features associated with the transition to laying eggs on land may therefore have been key in later enabling the evolution of terrestrial larvae and/or direct development. These features may include increased egg size and provisioning11,21, desiccation avoidance adaptations64, enhanced embryonic gas-exchange capabilities65, and parental care27,28. However, some of these characteristics may not be a requirement for live-bearing, in which neither the egg nor the developing embryo are exposed to the environment. Hence, in frogs, live-bearing seems to have evolved separately. The evolution of live-bearing in salamanders remains equivocal, evolving from fully aquatic or semi-terrestrial ancestry. In caecilians, the origin of live-bearing from direct development is strongly supported across methods.
Discordances in the evolution of live-bearing across clades merits further investigation. Overall, the evolution of live-bearing in vertebrates seems unlikely to conform to a single scenario66. For example, some live-bearing salamanders67 and frogs68 give birth to individuals in advanced larval stages. The modeling of live-bearing evolution may also be influenced by uncertainties in reproductive-mode coding and limited phylogenetic sampling of key species. For instance, there is some uncertainty around the reproductive modes of the cave salamander (Speleomantes sarrabusensis) that may be capable of bearing live young69 (but see ref. 70). There is also uncertainty about the reproductive mode of the closest living relatives of the live-bearing anuran genera Nectophrynoides and Nimbaphrynoides71. Furthermore, a possible third origin of live-bearing in frogs has been reported in the now extinct Eleutherodactylus jasperi, in a genus that is otherwise direct developing72.
Previous studies in frogs have found that direct development and terrestrial biphasic development may have evolved directly from fully aquatic ancestors20,21. The hypothesis-free BayesTraits analysis supports such direct transitions. Interestingly, we show that such a signal can also be obtained even if such transitions do not occur in the underlying state-transition model. When reconstructing ancestral states based on the best-fitting, semi-sequential corHMM model and estimating transition frequencies between nodes only (using a joint-estimation method), we recovered at least some direct transitions from fully aquatic development to direct development and terrestrial biphasic. However, these transitions did not occur when counting anagenetic transitions (through stochastic character mapping). We cannot rule out the occurrence of direct transitions, but observations of direct transitions may still involve intermediate, semi-terrestrial ancestors, as long as these occur rapidly and along a single branch21.
In support of this idea, we find that the transition rates and frequencies estimated for the loss of the larval stage (from semi-terrestrial to direct development) are among the highest of any state transitions in Anura and Caudata. The loss of the larval stage may thus have evolved rapidly through simple regulatory changes that are not easily captured by macroevolutionary methods. It is often a challenge for comparative analyses to reconstruct evolutionary patterns in developmental modes when evolutionary changes are rapid50. To understand how such seemingly complex changes in life-cycles can occur rapidly, we may need to gain a better understanding of their genomic and developmental causes. Developmental studies have found that direct development may evolve through heterochronic shifts in key developmental mechanisms, such as the shift in peak thyroid hormone levels during the embryonic period56,61,73,74,75. Similarly, viviparity in Salamandra salamandra may have evolved via accelerated development and heterochrony of the feeding and digestive systems67.
Reproductive modes and diversification rates
Our results challenge the idea that direct development or terrestrialization in anurans increase diversification rates22,23,24,25. To our knowledge, substantial support for this hypothesis has only been found for direct development in a single frog family (Rhacophoridae33). Across Anura, we find little evidence to support this claim. We do find heterogeneity in diversification rates within Anura that is attributable to unmeasured (hidden) trait states. Our results do not address what these hidden states are, but previous studies have shown that climate76,77,78,79, microhabitat79, and climatic-niche evolution78 impact anuran diversification.
In contrast, we found that salamander species with direct development have higher diversification rates. Direct development is also the most common reproductive mode among salamanders, unlike in frogs and caecilians. Interestingly, direct development has only evolved in a single family, Plethodontidae. This family includes the only salamanders that colonized the Neotropics, where they diversified extensively80,81. All tropical plethodontids have direct development, but there are also many temperate plethodontid lineages with direct development (e.g., Batrachoseps, Plethodon). The method we used ideally captures rate variation produced by unmeasured, hidden trait states (e.g., biogeography). However, the few origins of direct development in salamanders may make its direct effects on diversification difficult to disentangle from other clade-specific factors.
Intriguingly, we found that paedomorphism was associated with negative diversification rates (extinction rates higher than speciation rates). This is consistent with the numerous repeated origins of paedomorphosis (the most common reproductive-mode transition in Caudata), but the low number of extant paedomorphic species. There is some support for a negative relationship between diversification rates and paedomorphosis among salamander families36. Similarly, an analysis of spelerpine plethodontids37 found that paedomorphosis increased speciation rates but increased extinction rates even more, leading to decreased net diversification rates. We also find this same speciation-extinction dynamic resulting in negative diversification rates here (Supplementary Data 2). We therefore strongly support the hypothesis that paedomorphosis can decrease diversification.
Unlike Anura and Caudata, we estimated diversification rates to be low and constant across Gymnophiona. Diversification rates in Gymnophiona may also be largely constant over time82. Why this may be remains unclear. There is a trend toward lower diversification rates in fossorial frogs79 (but not significant) and fossorial squamates83,84. Fossoriality is common in caecilians4, and might explain their relatively low and constant diversification rates.
In conclusion, here we present a large-scale comparison of evolution of life cycles and reproductive modes among the three major amphibian clades. We found that transition rates among states are largely decoupled from their species richness. Specifically, we found that in all three groups, the most frequent transitions were to states that are relatively uncommon in that group (live-bearing in caecilians, paedomorphosis in salamanders, semi-ter...



Comments
Post a Comment